Activation of Nerve-cell Structures in Mice and Astronomists: Detection of Sound Waves and its Implications for Astronomers
150 years after they were discovered, researchers have identified how specific nerve-cell structures on the penis and clitoris are activated. There was no known about the role of these structures in sex or how they work. When exposed to low- Frequency vibrations, a team of mice were shown to have Sexual behaviours like erections, and both male and females were activated as a result. The researchers hope that this work could help uncover the neurological basis underlying certain sexual dysfunctions.
Astronomers have a difficult time figuring out the identity of a mysterious object and how a CRISPR gene editing could make rice plants more water efficient.
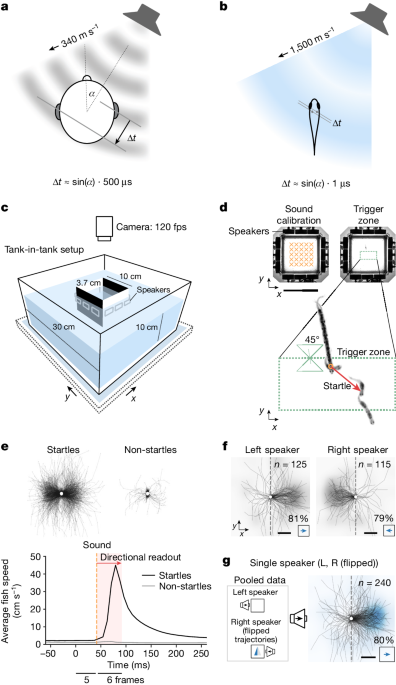
It has been thought that fish can identify the direction a sound is coming from but it is a question that has puzzled scientists for years. Now using a specialist set-up, a team of researchers have demonstrated that some fish can independently detect two components of a sound wave — pressure and particle motion — and combine this information to identify where a sound comes from.
Tune in with Nature Podcast: Detecting the motion of the inner structures of a fish with a laser-scanning vibrometry
Never miss an episode. You can subscribe to the Nature Podcast through the following methods: applepods, sebastian,youtube music or your favourite app. An RSS feed for the Nature Podcast
is available too.
To analyse the motion of the inner structures of the fish, we used Matlab 2019b and a particle image velocimetry toolbox PIVlab80, originally developed to characterize the motion of flowing particles for fluid mechanics. The cross-correlating subregions are used to assess the particle displacement. The contrast of the images was improved before the displacement analysis and the results werecurated in post-processing by removing outliers.
To reconstruct the relative phases of a motion, we needed to measure each phase under more than two different phases. As noise can influence this measurement, we used four phase steps here, ensuring proper phase reconstruction while keeping acquisition sessions reasonably short.
The principle of the laser-scanning vibrometric measurement is illustrated in Fig. The data in 3b and 10b is extended. The samples was stimulated with an acoustic wave at a certain frequencies and imaged with a laser-scanning microscope. 10b(ii)).
Source: The mechanism for directional hearing in fish
Micro-CT scans of line-ablated wild-type Drosophila cerebrum fish using a laser-scanning two-photon microscope
The fish were injected with a dose of 120 g l1 per fish. They were placed on a preformed agarose mould which allowed the gill protects to move freely, and immobilized with 2% low-melting-point agarose. A flow of aerated aquarium water flowed into their mouth using a glass capillary.
The confocal microscope was based on a custom-built laser scanning two-photon microscope. The illumination source was a Ti:sapphire laser. Before entering a laser-scanning two-photon microscope, the beam passed through a 90:10 beam splitter (90% reflection, 10% transmission). The light back-scattered by the fish inner structures was descanned, reflected by the 90:10 beam splitter, and then focused by a lens (f = 50 mm) into a single-mode fibre (core diameter: 25 µm, numerical aperture: 0.1) acting as a confocal pinhole. The microscope was controlled by custom-written software (https://github.com/danionella/lsmaq).
There are 65 fish that are treated and the data stem from them. We show the number of fish that responded with a single startle. The same experiment, also comprising 12 sound configurations, was repeated with 74 lateral line-ablated fish (Extended Data Figs. 7 and 9; 2013 playbacks, 910 startles, 45% startles). A third sound playback experiment was carried out in the dark in 43 untreated fish, testing a subset of 4 sound configurations (Extended Data Fig. 12b).
A 12-month-old male wild-type D. cerebrum was euthanized by ice shock and fixed with 4% paraformaldehyde in phosphate-buffered saline (PBS) at 4 °C overnight. The fish was washed in PBS for 15 min and stained with 5%Sigma Aldrich solution in PBS at 4 C the next day. After staining, the fish was washed in PBS and then embedded in 1% PBS buffer inside a cryo tube. The micro-CT scan was carried out at the ANATOMIX beamline at SOLEIL synchrotron by XPLORAYTION. The sample was placed into a 40-keV polychromatic (white) X-ray beam. A scan consisted of 3,200 projections collected at about ×10 optical magnification by a digital camera (Orca Flash 4.0 V2) with a sensor pixel size of 6.5 µm at 150 ms exposure time, yielding an effective pixel size of 0.6485 µm. The registered data were binned to 1.2970 µm voxel size. Key structures of the hearing apparatus were manually segmented. 3D Slicer76 was used and then used to convert planes into a 3D model. FIJI ImageJ (v1.5)78 was used to convert between different file types. They turned the segments into mesh grids for cleaning and rendering.
There’s a bias per fish. In the analysis of bias across trials and fish, theoretically, all trials could stem from one performing animal (not of concern here; Extended Data Fig. 5a,b). To complement this measure, we also quantified the directional bias per fish. We had 12 sound configurations in each experiment and startles averaged to a total of about 22 startles per fish in an experiment; hence, a meaningful per-fish bias could be computed only on pooled sound configurations and for fish with many startles. To estimate the directional bias of individual fish, we filtered for fish that had ≥10 startles in both the single-speaker condition (pooled over 4 stimuli) and the trick condition (pooled over 4 stimuli; Extended Data Fig. 6c,d. Although the value reflects directional behaviour in the population and estimates fish-to-fish variability, it selects for fish that trigger many playbacks and startle often.
The swimbehaviour of D. cerebrum was tracked with a pose. The fish were recorded over a period of nine frames with the skeleton of seven equidistant points along their body segments and two additional points for each eye. The ‘single-animal’ model was used for training. Data availability can be found at the G-Node repository, which has the model parameters and trained model.
Each fish was tested once, and one fish was tested at a time. In the first couple of minutes of the recording, a 10 cm 10 cm plate was put into the inner tank to match the sound grid to the video frame. Three minutes after placing the fish in the inner tank, playbacks were triggered for 45 min.
The LAGeSo approved all animal experiments that followed the Berlin state, German federal and European Union animal welfare regulations. D. cerebrum were kept in commercial zebrafish aquaria (Tecniplast) with the following water parameters: pH 7.3, conductivity 350 µS cm−1, temperature 27 °C. We used male and female adult fish between 4 and 11 months of age.
Source: The mechanism for directional hearing in fish
Particle Acceleration at a Distance r(r,t) to a Sound Monopole with Discrete Signals from Different Phases of the Waveform
To compute the particle acceleration ({a}{r}(r,t)) at a distance (r) to a sound monopole with pressure (p(r,t)) for discrete signals of arbitrary waveform, we applied this equation separately for each Fourier component. Given a pressure waveform ({{{\bf{p}}}{n}}\,:={p}{0}{,p}{1},\cdots ,{p}{N-1}) with (N) samples ({p}{n}), spaced at (T=1/{sr}) with sample rate ({sr}), the particle acceleration ({{{\bf{a}}}{n}}\,:={a}{0}{,a}{1},\cdots ,{a}{N-1}) that would be observed at a distance (r={r}{0}) from a sound monopole was calculated by carrying out the discrete Fourier transform ({{{\bf{P}}}{l}}\,:={P}{0}{,P}{1},\cdots ,{P}_{N-1})
The additional constraints on the scanning parameters were set by this. We used ({f}{{\rm{scan}}}=800\,{\rm{Hz}}) and ({f}{{\rm{stim}}}=\mathrm{1,000}\,{\rm{Hz}}) for the data presented in Fig. 3 and Extended Data Fig. 11.
To ensure repeatable measurements, the acoustic stimulation and the galvanometric scanning mirrors were synchronized so that each pixel was recorded at a known sound phase. This was achieved by triggering the sound generation on each single frame scan trigger.
We observed that D. cerebrum startle when we drop a cylindrical piece of rubber into the water. The high-pass was used to get the pressure for the sound at 100 Hz, and we used a 12-ms fragment to serve as our template for the pressure of the speaker signals. The target pressure amplitude was set to a peak sound pressure level of 167 dB (referenced to 1 µPa) by rescaling this pressure waveform accordingly. This amplitude was loud enough to elicit startles reliably and still supported by our small 2.7-cm-diameter speakers. The first peak was about 0.664 ms and the centre Frequency was about 780-0, 000 hertz. The pressure and monopole theory were used to calculate the target horizontal particle acceleration.
and with frequency f, (\omega =2{\rm{\pi }}f), wavenumber (k=2{\rm{\pi }}/\lambda ), wavelength (\lambda ) and speed of sound ({c}=\,\lambda f).
The radial particle velocity can be decays with or without the distance in the near field.
By contrast, particle acceleration—the temporal derivative of particle velocity—decays quadratically with distance for nearby sounds ((r\ll 1), limit independent of frequency):
The particle acceleration was measured with a triaxial ICP and NI-9 sound and vibration module that was acquired by PCB Piezotronics. 1d The hydrophone has an acceleration sensor that is moved across the grid positions by repeating the same sound.
The indirect method was used in all experiments to measure x and y acceleration. The particle acceleration sensor still proved useful in measuring the vertical z acceleration in our setup.
Whereas hydrophones are manufactured and calibrated for underwater use, the particle acceleration sensor is not made to measure particle acceleration underwater and is meant to be glued onto the vibrating object. Owing to an acoustic impedance mismatch between metal and water, we expected the PCB sensor to underestimate particle acceleration.
We compared x and y acceleration waveforms for both measurement methods and found that the acceleration waveforms acquired through the direct method match the waveforms acquired through the indirect method after multiplication by a factor of about 2.4. The close match in rescaled waveforms confirms the validity of the gradient approximation in the indirect method.
Source: The mechanism for directional hearing in fish
Rescaling of a Nonlinear Speaker Signal Wave in a Strongly Coupled Electromagnetic Field: Supplementary Table 2
To increase robustness of the solutions and to avoid speakers canceling themselves, speaker signal waves were forced to be similar to the target. The system of equations was solved with a least-square solver. The absolute components of the target pressure pattern were rescaled as a bound (B_i,l).
Past s_i ast mathopsum.
in which (\gamma ) is fixed and scales pressure to voltage and ({\alpha }_{i}) is a rescaling parameter set independently for each speaker to give additional control over active speakers. The values are listed in Supplementary Table 2.