Network analysis of diet- and body-weight-adjusted correlations across a lifespan-based study in Bar Harbor, Maine
Gary was associated with the study at the Jackson Laboratory in Bar Harbor, Maine. But they do not lead to lifespan extension.
The results drive home the intricate nature of the body’s reaction to restriction. “It’s revelatory about the complexity of this intervention,” says James Nelson, a biogerontologist at the University of Texas Health Science Center in San Antonio.
The study was published today in Nature by Churchill and his co-authors, including scientists at Calico Life Sciences in South San Francisco, California, the anti-ageing focused biotech company that funded the study.
Daniel Belsky, an epidemiologist who studies ageing at the Columbia University Mailman School of Public Health in New York City, cautions against over-extrapolating from mice to humans. He acknowledges that the study adds to the belief that lifespan and healthspan are different things.
In M2, we looked at the regression coefficients of the trait term in relation to the lifespan. As the traits are standardized, the regression coefficients are equivalent to diet- and body-weight-adjusted partial correlations (designated as r) and can be compared across traits. Moreover, we stratified the data to estimate diet-specific body-weight-adjusted partial correlations that are shown in the scatter plot panels (Fig. 2f).
We performed a multivariate network analysis to decompose the effects of DR on lifespan across measured traits. An empirical covariance matrix was estimated using the nonparanormal SKEPTIC estimator derived from the pairwise Kendall–Tau correlation using pairwise-complete data. The estimated covariance matrix was projected to the nearest positive definite matrix by truncating eigenvectors with negative eigenvalues. A sparse low-rank graphical model was used by the ggLASSO Python package. The result was normalized to obtain an inferred partial correlation matrix, which we use as our phenotype network for downstream analysis.
BW is body weight at the day closest to T’s collection date, it is the percentage of life lived as of T collection date, and s is the smoothing parameters. Each mouse had multiple datapoints across the T collection date. This clustering was accounted for with a random intercept for ID, specified as (1|ID) above. We used our hypothesis tests to find out how much trait sensitivity model 1 has to body weight. Using the models specified above and a conservative false-discovery rate (FDR < 0.01, one-step Benjamini–Hochberg method), we therefore identified traits that responded additively to body weight, traits that responded additively to diet, traits that responded additively to PLL (scaled age) and traits that responded interactively to diet and PLL. Health, metabolism, haematology and immune were the traits considered to have health. The number of traits with significant associations with body weight are shown in the bar plots. In place of PLL we repeated the analysis with age. We applied FDR adjustment to each test across traits and timepoints (Benjamini–Hochberg FDR method).
When testing, the body weight is at the time of testing, not the last preintervention body weight. For body composition and change-in-body weight traits, we did not include the body weight terms. Before model fitting the rank normal scores of all continuous variables were transformed. We performed likelihood ratio tests for the diet and body weight adjusted association (M2 versus M1) and for the diet × trait interaction (M3 versus M2). We applied a FDR adjustment to each test across traits and timepoints (one-step Benjamini–Hochberg method). Traits were categorized and significant (FDR < 0.01) results were tabulated as above.
Climber was used to track animals and provide a stable repository for primary data collection. Data was frequently reviewed by a statistical analyst. Equipment miscalibration, mislabelled animals, and technically impossible values are some of the problems found in the initial data quality control. If we could not manually correct these using laboratory records, they were removed. Body temperature and body weight were explored for outliers. There were corrected Quantitative traits other than body weights. We use a fully random-effects model conditioning on diet, body weight and age to quantify the effects. The values were adjusted by subtracting the coefficients. The lifespan data is presented in months for easy interpretation. The main text contains a statistical significance for extremely small P values of P 2.2 1016 and non-truncated P values are given in the Supplementary Information. All analyses were performed using R v.4.2.2 and RStudio v.2022.12.0+353. Data analysis scripts are available to be used.
Research staff evaluated mice for a variety of clinical symptomology such as responsiveness to stimuli, being able to eat or drink, scurvy, wounds of aggression and abdominal distention. If mice were found to meet the criteria for observation, they were contacted by the veterinary staff. If the clinical team determined a mouse to be palpably hypothermic and unresponsive, unable to eat or drink, and/or met protocol criteria for severe dermatitis, tumours and/or fight wounds, pre-emptive euthanasia was performed to prevent suffering; otherwise, the veterinary staff provided treatment. The death of a mouse is represented as a death in the survival curves. The euthanized mice were treated as censors and had a total of 13 events recorded.
Free-wheel-running data were collected at around 44, 96 and 144 weeks of age. Mice were individually housed for a minimum of 36 h in a special cage suited to house the Med Associate low profile running wheel with a wireless transmitter. The food hopper was removed to allow for seamless movement of the wheel, and food was placed onto the cage floor. The 15.5-cm-diameter plastic wheel sits at an angle on an electronic base, which tracks the revolutions. The battery-powered base allows for continuous monitoring of data, which is then wirelessly transmitted, in 30 s intervals, to a local computer.
Cages were prepared by cutting a piece of cosmos blotting paper, 360 gsm, to standard duplex cage dimensions. The paper was taped to the back of the cage after shavings were removed. The water was removed to prevent it from leaking onto the paper during the test. The cages were prepared for mice to be housed in. At the end of the trial, the mice were returned to their original housing units, and papers were removed and dried for 2–4 h, before being individually bagged. Papers were shipped to Beth Israel Deaconess Medical Center, where they were scanned with ultraviolet light to image and quantify the void spots.
We used the Ugo–Basile rotarod, which has five lanes evenly spaced along a motorized horizontal rotating rod, allowing for up to five mice to be tested simultaneously. There is a platform below each lane that records the height of each mouse as it falls. At the beginning of the session, the mice were placed on a rod which began rotating at 4 revolutions per minute and continued for a maximum of 40 revolutions per minute. We reported the mean latency (time to fall) and the slope of latencies across trials, as well as the number of trials with no falls and number of trials with immediate falls. trials were recorded if a mouse didn’t cooperate with the test.
Startle response was measured in rodents using automated startle chambers, in which a mouse was placed in a clear, acrylic tube attached to a highly sensitive platform that is calibrated to track their startle reflex while being exposed to a series of stimuli at varying decibels and times. Mice were initially exposed to white noise from an overhead speaker, which transitions to a series of randomized, computer-generated stimuli ranging in volume from 70 to 120 decibels at 40 ms in duration and an interval of 9–22 s. The test takes about 30 minutes.
Ultrasonography of mice: Determination of circulating immune cell subsets using flow cytometry and blood reagents in outbred DO mice
Ultrasonography was performed using the VisualSonics (VSI) Vevo 770/2100 high-frequency ultrasound system with 30 and 40 MHz probes. It uses a probe to measure blood flow rates and volumes.
At the flow cytometry blood collections, mice had to be fasted for 4 h and measured using the One Touch Ultra 2 meter from LifeScan along with One Touch Ultra test strips. At each of the CBC blood collections at 24, 71 and 122 weeks, non-fasted glucose was measured using the glucose meter.
Owing to the outbred nature of these mice, flow cytometry markers were limited, and T cell subsets were generally assigned as naive and non-naive by the presence of CD62L and CD44 (immune cell subtype designations are shown in Supplementary Table 7). Memory T cells that accumulate after immune responses are enumerated and may represent NKG2D-positive cells. Owing to limitations in flow cytometry markers that identify NK cells and their subsets in the mouse strains contributing to the outbred DO mouse line, NK cells were defined as non-T non-B lymphocytes expressing NKG2D. Within this population, CD11c and CD11b were used to generally define maturation subsets. The expression of CD11b and CD11c is marked by more mature NK cells.
Flow cytometry can be used to determine the frequencies of circulating immune cell subsets. Analysis was performed before the start of dietary interventions at 5 months, then at 16 and 24 months of age. These timepoints corresponded to 11 and 19 months of dietary intervention. Red blood cells in PBL samples were lysed and the samples were washed in FACS buffer (Mitenyi, 130-091-222). Cells were resuspended in 25 μl FACS buffer with 0.5% BSA (Miltenyi, 130-091-222 with 130-091-376). Antibodies including Fc block (2.42, Leinco Technologies) were added and incubated for 30 min at 4 °C. After the cells were washed, the DAPI was added to the LSriI system. The antibody cocktail contained CD11c FITC, N418 (35-0114-U100, Tonbo Biosciences, 1:100); NKG2D (CD314) PE, CX5 (558403, BD Biosciences, 1:80); CD3e PE-CF594, 145-2C11 (562286, BD Biosciences, 1:40); CD19 BB700, 1D3 (566411, BD Biosciences, 1:40); CD62L PE-Cy7, MEL-14, (60-0621-U100, Tonbo Biosciences, 1:100); CD25 APC, PC61 (102012, BioLegend, 1:80); CD44 APC-Cy7, IM7 (25-0441-U100, Tonbo Biosciences, 1:40); Ly6G BV421, 1A8, (562737, BD Biosciences, 1:80); CD4 BV570, RM4-5 (100542, BioLegend, 1:40); CD11b BV650, M1/70 (563402, BD Biosciences, 1:160); CD45R/B220 BUV496 (RA3-6B2, 564662, BD Biosciences, 1:20); Fc Block, 2.4G2 (C247, Leinco Technologies, 1:100).
Source: Dietary restriction impacts health and lifespan of genetically diverse mice
Bone density and composition measurements using the LUNAR PIXImus II densitometer and 3D NMR to monitor the effects of fasting in female DO mice
We used the LUNAR PIXImus II densitometer to collect bone density and body composition. mice were put onto a disposable plastic tray that was then placed on the exposure platform of the PIXImus The process to acquire a single scan lasts approximately 4 min. There were three measurement taken at 96 weeks, 44 weeks and 144 weeks of age.
The modified version of the clinically relevant FI38 was used by us to calculate the average of 31 age-associated discrepancies and health decline indicators. The missing deficit, mild deficit, and severe deficit were all scored on a scale of 0, 0.5, or 1, where 0 indicated the absence of the deficit. Measurements were taken at the baseline (5 months) and were repeated approximately every 6 months. Simple averaging yielded a raw FI score of between 0 and 1 for each mouse. Frailty scores were adjusted by estimating batch, coat colour and experimenter effects as random factors that were subtracted from raw frailty score values before statistical analysis.
The 959 mice had over 100,000 values collected over the course of their lives. Body weights were looked at after local regression fitting within mouse.
To obtain an accurate estimate of food intake and changes in body weight in response to weekly fasting cycles, we set up an independent cohort of 160 female DO mice. Mice were placed on the same DR protocols as in the main study. Food was weighed daily for a period of 1 week when mice were 30, 36 and 43 weeks of age. The food intake data was normalized to units of g per mouse per day and presented as daily and weekly averages. At 43 and 45weeks of age, body composition was determined with the use of an instrument that can be used in small- animal studies and a 5-gauss magnet. NMR data were used to detect changes in body weight and composition before and after fasting. Pre-fasting values of body weight, lean mass, fat mass, and adiposity was plotted with the difference between before and after fast.
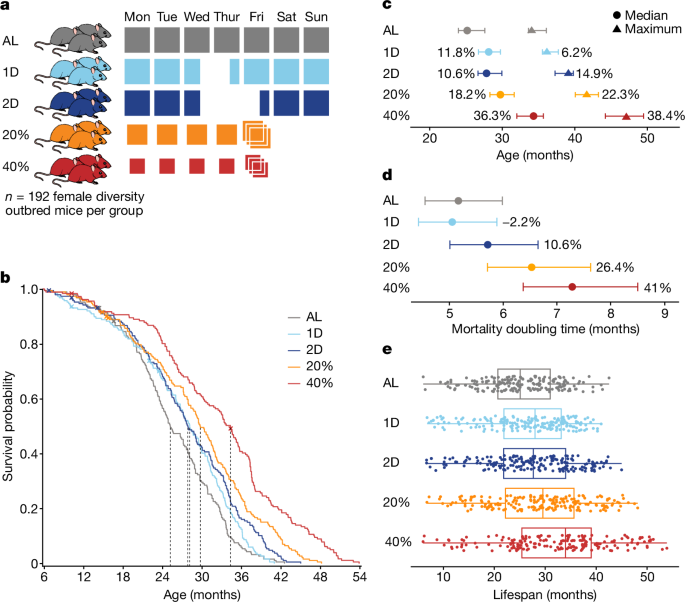
DR was implemented by controlling the timing and amount of food provided to mice. Feeding schedules for DR were started at 6 months of age. There was a standard mouse chow diet that all mice were fed. The group was provided unlimited access to food and water. The IF mice were provided unlimited access to food and water. On Wednesday of each week at 15:00, IF mice were placed into clean cages and food was withheld for the next 24 or 48 h for the 1D and 2D groups, respectively. CR mice were provided with unlimited access to water and measured amounts of food daily at around 15:00, 2.75 g per mouse per day for 20% CR and 2.06 g per mouse per day for 40% CR. These amounts were based on AL consumption of 3.43 g per mouse per day that we estimated based on historical feeding data from DO mice. The mice were first subjected to 20% CR for 2 weeks, then 30% CR for another 2 weeks before transitioning to 40% CR. The mice were first acclimatized to the 1D IF regimen for 2 weeks. The mice were kept in pens with eight mice per pen. Co-housing is standard practice for CR studies39; competition for food was minimized by placing food directly into the bottom of the cage, allowing individual mice to ‘grab’ a pellet and isolate while they eat. The 40% CR mice were provided with a 3 day ration of food on Friday and followed by a period of food deprivation of about 1 day, which was comparable to the intensive fasts of the IF. The 15:00 feeding time closely approximates the circadian alignment of feeding, starting just before the beginning of the dark cycle, which is the normal active and feeding time of day for mice. This timing has been shown to maximize lifespan extension in mice subjected to 30% CR8. The Friday triple feeding is used in other studies but there is no direct assessment of the health impacts.